La materia che ci accomuna è molto ampia, e certi processi fisiologici, per poter essere compresi davvero, oltre che lo studio della letteratura scientifica richiedono anche delle profonde riflessioni. Sono quelle stesse competenze che abbiamo acquisito negli anni a permetterci una migliore organizzazione delle idee, le quali, senza un solido rudimento in materia, non potrebbero tuttavia essere razionalizzate.
Anche i confronti con i colleghi, magari dotati di competenze a noi complementari, possono dimostrarsi validi spunti di riflessione. È il caso dell’ultimo scambio avvenuto tra me ed Emanuele Giordano, con il quale collaboro da ormai diversi anni nel progetto scientifico-divulgativo Mission Evolution Sport.
L’argomento in oggetto riguardava la partecipazione delle fibre a contrazione rapida in presenza di carichi medio-bassi (nella fattispecie 50% di 1RM) e velocità di contrazione “super slow” a tensione continua.
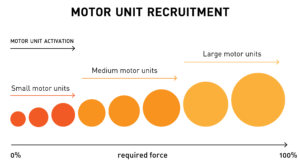
Il nocciolo del dibattito era incentrato sul fatto che le fibre rapide (IIb, per convenzione IIx) fossero coinvolte o meno nelle fasi iniziali dell’attività muscolare. Personalmente, guardando al principio di Henneman, sostengo che la partecipazione di fibre rapide sia relativa all’intensità di carico (% di 1RM, vedi schema allegato) ed alla velocità di contrazione, suggerendo pertanto l’esclusione, per lo meno in una prima fase (e poi vedremo perché), delle fibre IIx. Giordano invece cita gli ultimi studi elettromiografici condotti sul movimento che, andando in contrasto con la stessa legge di Henneman, indicano un intervento pressoché immediato (per motivi circostanziali) delle unità contrattili più rapide. Lo stesso a riguardo riporta una ricca bibliografia nel testo di Alberti, Garuffi e Nicola Silvaggi.
Malgrado non abbia avuto modo di visionare tali pubblicazioni, cercherò di approfondire il concetto con l’ausilio di un’indagine osservazionale e logico-deduttiva. Partiamo innanzitutto dal presupposto che la capacità d’ipertrofizzare sia prerogativa delle fibre a contrazione rapida ed intermedia, prevedibilmente in maniera più favorevole alla prima categoria. Teniamo pure in considerazione che, a prescindere dalla velocità di contrazione adottata nonché dal minore o maggiore sforzo raggiunto, l’utilizzo di alte intensità di carico (>80%) ne obbligherà comunque l’attivazione.
La dimostrazione di quanto appena detto è il fatto che powerlifter e weightlifter, lavorando con carichi mediamente elevati, pur non conducendo abitualmente i set ad esaurimento, mostrano comunque notevoli sviluppi ipertrofici. Il bodybuilder, che invece sfrutta ordinariamente la pratica del cedimento muscolare, si concede anche ai carichi più “piccoli” (in termini di intensità di carico ovviamente), innescando così gli importanti cambiamenti metabolici alla base dei consistenti adattamenti ipertrofici che noi tutti possiamo apprezzare (la cui maggiore rotondità può essere individuata nella componente sarcoplasmatica, anch’essa legata all’enfasi sul sistema energetico).
Ora, se dovessimo soffermarci all’essenza stessa del risultato, potrebbe apparire scontato l’impiego delle fibre a rapida contrazione sia in una che l’altra tipologia di atleti; ciò che invece rimarrebbe ostico sarebbe l’entità dei meccanismi di reclutamento innescati. Ed è proprio questo aspetto che volevo approfondire.
Probabilmente la legge di Henneman non godrà di valore assoluto, ma tuttavia si sposa perfettamente con la teoria del “tutto o del nulla” (una fibra o è attiva o non lo è), alimentando dunque la tesi di come una maggior forza possa essere prodotta nient’altro che da un numero maggiore di unità motorie sinergicamente attive. Ciò fa pensare che gli schemi di attivazione muscolare siano soggetti ad un’organizzazione neurale ben precisa, e sulla base di questa osservazione si dovrebbe considerare l’esistenza di motivazioni più valide affinché il CNS assegni un lavoro inappropriato alla categoria delle “fast twitch” (leggi II x). Il mio pensiero va all’ambiente metabolico.
Mi spiego meglio: quando si effettua un lavoro altamente selettivo nonché a tensione continua, se l’entità del carico è abbastanza alta, durante l’esercizio si andrà creando una forte compressione vascolare la cui cosa impedirà il momentaneo afflusso di sangue. Questo fenomeno, altrimenti conosciuto come “blood flow restriction” (BFR non da laccio emostatico in questo caso), creerà presto o tardi una certa ipossia tissutale, ergo un ambiente fortemente anaerobico. Ecco il primo dei motivi che obbligherà le fibre a rapida contrazione ad entrare in gioco, poiché sarebbero proprio loro le uniche in grado di lavorare in assenza di ossigeno (non essendo dotate di mitocondri, hanno un metabolismo prettamente glicolitico).
Il secondo motivo sarà individuabile nel progressivo esaurimento delle prime reclute (che a mio parere ci rimanda alle fibre di categoria I e IIa), in quanto la tensione continua in assenza di sufficienti spazi di recupero, saprà come determinarne il progressivo esaurimento. In pratica man mano che le unità motore più lente (più pertinenti ai sopracitati parametri di lavoro) si esauriscono, entrano in gioco le successive; l’ambiente metabolico non farà altro che accelerare il fenomeno, ma perché ciò accada dovranno trascorrere diverse decine di secondi dall’inizio del lavoro, ed in super slow i tempi si dilatano ulteriormente.
Quello che possiamo evincere da un’analisi di questo tipo, è l’effettiva funzione del parametro “intensità di sforzo” (ripetizioni effettuate vs ripetizioni fattibili); in soldoni, se la % di carico è ridotta (50-60% di 1 RM) e la velocità di contrazione lenta, sarà del tutto plausibile il mancato ingaggio delle fast twitch (e dunque una minora risposta ipertrofica), pertanto condurre il set a cedimento potrebbe mostrarsi come l’unica garanzia per l’attivazione delle fibre bianche (ovvero fast twitch, o II b o IIx).
Ma c’è un però. Con l’aumento esponenziale del rateo di scissione di ATP, si va incontro ad un precoce calo dei livelli energetici, con conseguenze che si ripercuoteranno sia sul volume di lavoro potenzialmente cumulabile, nonché sulla concentrazione di ioni H dentro la cellula. Quando il primo potrebbe pregiudicare la grandezza dello stress meccanico (proporzionale alla quantità di lavoro prodotto), il secondo, oltre ad essere motivo di crescita dello stress metabolico, potrebbe anche sconvolgere l’intero equilibrio osmotico della stessa cellula muscolare.
Perché accade ciò?
Per motivi legati al bilanciamento anionico. La carenza di ATP, l’inibizione dall’attività chinasica indotta dall’aumentata idrogenia intracellulare, blocca l’attività della pompa sodio/potassio; ciò porta ad accumulare Na+ intracellulare ed escretare potassio, senza permettere a quest’ultimo di essere riassorbito.
Risultato?
Aumento dell’ECW (extra cellular water) a scapito dell’ICW (intra cellular water).
A tutto questo aggiungiamoci una potenziale inibizione di M-tor dovuta ad un proporzionale aumento di AMPK, essendo quest’ultimo una sorta di termometro energetico della miocellula. È vero che l’aumentato rateo di scissione di ATP promosso dall’esercizio possa essere motivo di attivazione dell’isoforma alfa 2 di AMPK (in teoria non inibente M-tor), ma dal punto di vista meramente osservazionale, spingendo troppo sullo stress metabolico, non posso dire di aver mai registrato importanti cambiamenti ipertrofici. Rimango pertanto dell’opinione, che nel soggetto natural debba essere rispettato il rapporto d’equilibrio vigente tra i principali stressor, di cui ho ampiamente discusso in diversi post sulla pagina ufficiale di Facebook.